the metabolic rift: how global modes of material exchange disrupt local biogeochemical cycles.
As the culmination of the Harvard GSD MDes program, each student is assembled into a small team to complete an Open Project Thesis motivated by a particular instructor’s interests. Our class, Metabolic Rift, Shift and Gift was taught by Kiel Moe, an architect who pioneered ecological systems analysis in the construction of buildings. To bring the metabolic rift into focus, we aimed our studies at one particular species in our own local ecology: the Eastern Hemlock. We spent the semester studying the epistemic, ontological, material, and ecological relations that produce the Hemlock as an entity in a cultural assemblage and developed individual “gifts” to repair the severed human-ecological relationship at the base of this rift.
The contemporary Eastern Hemlock has existed for at least 16,000 years. Since time immemorial, various Indigenous peoples—including the Abanaki, Seneca and Mi’kmaq—have understood the Eastern Hemlock as an agentic relative who provided them with life-sustaining gifts: pine needles for medicinal tea, bark for dying fabric, boughs for bedding material, and a shaded understory to attract deer. In the Abenaki language, the name for hemlock is alnizedi, where alni denotes familiarity in interpersonal relations and zedi denotes the structure of coniferous trees. With alnizedi perceived as an extension of the social sphere, the Abenaki and Hemlock population were historically distributed over highly coinciding geographic ranges. Thus, their metabolic modes of material exchange enabled them to reproduce as a continuous socio-ecological system over time.
The ontological extension of the Self determines the bounds over which metabolism is optimized for the reproduction of a socio-ecological system. Neoplastic contractions of the Self render non-human relatives as resources. This is the perceptual pretext from which the metabolic rift emerges, as the unit of reproduction is constrained down to Anthropos.
In the wake of European colonization, it was found that the bark could be used for the production of leather goods and low-fidelity architectural materials. The ensuing clear-cutting of hemlock stands fueled the accumulation of capital in urban centers at the expense of the tree’s ability to regulate local microclimates, degrading the regenerative capacity of the Northeast’s forests over time. Eventually, cheaper inputs were discovered in leather production and the Hemlock’s primary use-value shifted towards the input for newspaper and gift wrap.
In the 1950s, the Wooly Adelgid was accidentally introduced into Eastern Hemlock forests and has produced catastrophic population decline. While there have been discoveries of locally resilient hemlock subspecies, the future longevity of the Eastern Hemlock remains uncertain. It is from this context where we set out to repair the rift economy through small acts of a gift economy.



Image Credits: Justin Booz
The Gift:
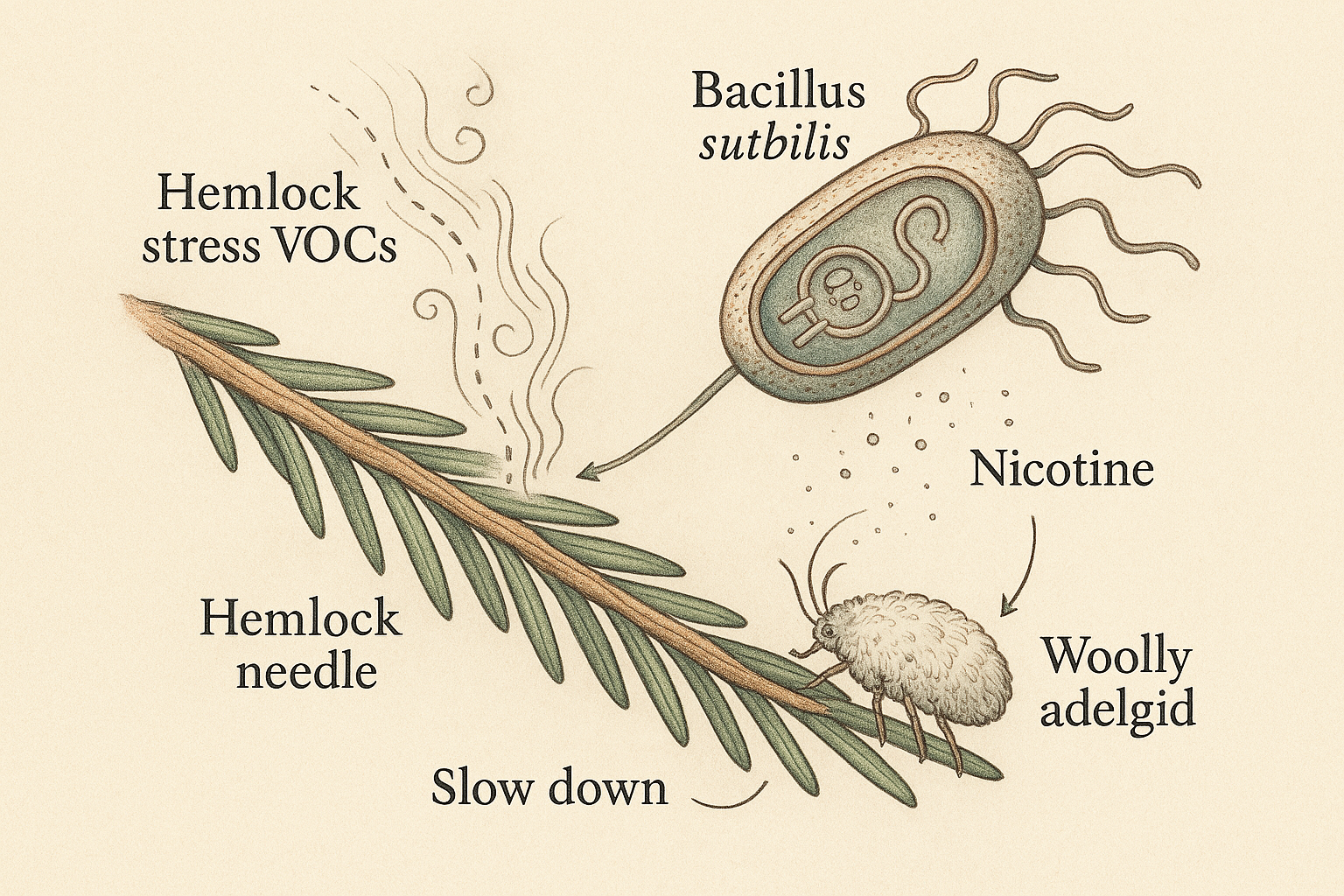
Forest ecosystems utilize mechanisms of stress-sharing to optimize system-level goals. Through airborne signaling of volatile organic compounds and subterranean mycorrhizal communication, individual trees are able to share stress signals with each other to prepare for dangerous predators, coordinate seed dropping, and distribute water to saplings. The wooly adelgid cannot “perceive” these stress signals which would otherwise coordinate their behavior toward system-level homeostasis. Therefore, I sought ways to bring the wooly adelgid into this semiotic regime through using a familiar bacterial interlocutor as a translator of arboreal stress signals:
Bacillus subtilis.
After walking through the Arnold Arboretum on the outskirts of Boston, it became clear that what Hemlock needs most at this moment is medicine. Given my background in bioengineering, I thought I might be able to help.

Bacillus subtilis is a relatively docile, commensal bacteria which lives all across the earth’s surface—including the needles of Hemlock trees. Given that it naturally engulfs DNA from the environment, it is also a workhorse for synthetic biology. Would it be possible to work with B. subtilis as a translator to convert Hemlock stress signals into a format that the Wooly Adelgid can understand? Might this shift wooly adelgid population behavior to more sustainable levels of consumption?
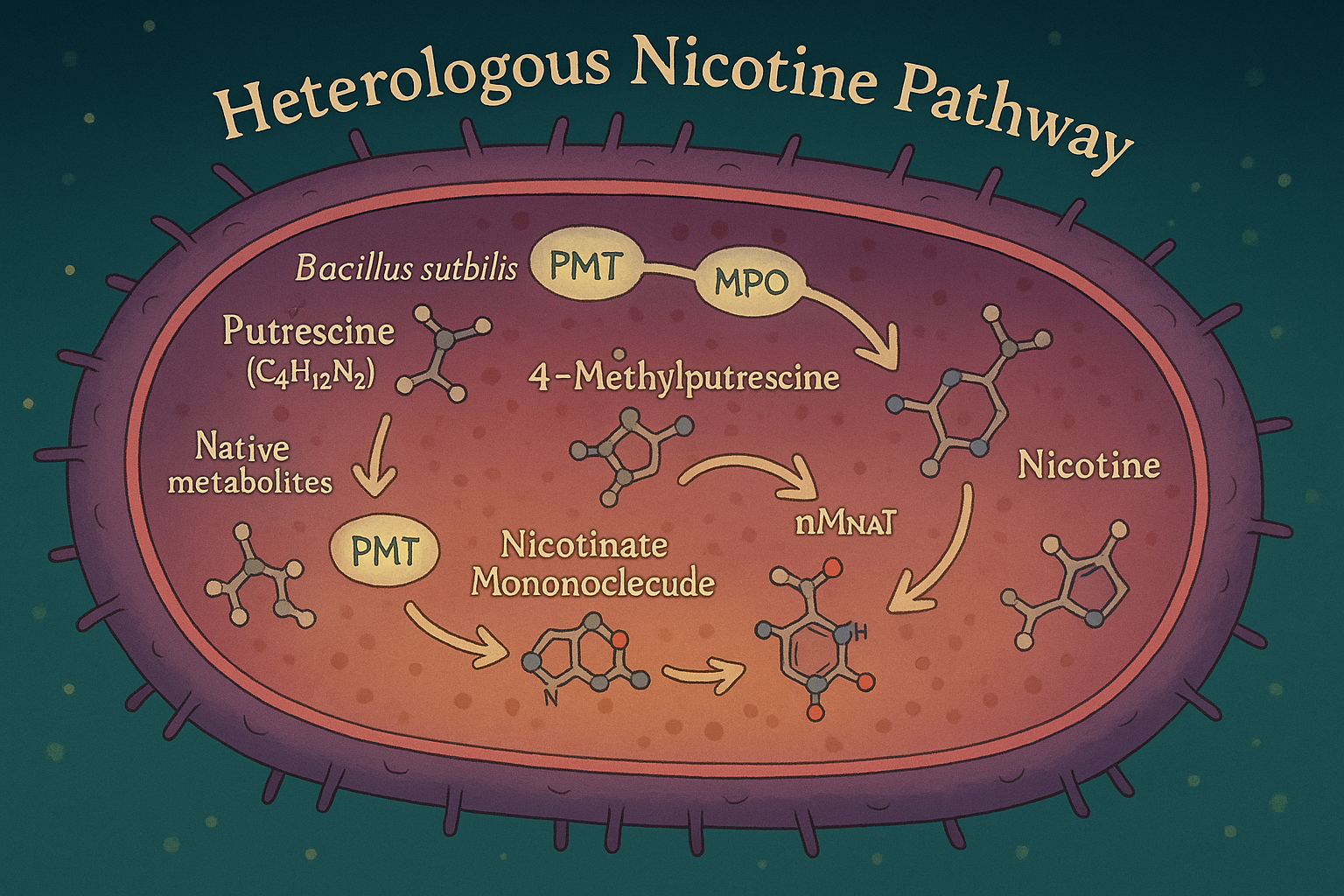
I began by investigating the molecular signals which cause stress for the wooly adelgid. One potent molecule, often used in pesticides, is nicotine. Thus, I designed metabolic pathways for Bacillus subtilis to start producing nicotine from native metabolites and tobacco plant genes.

To fix the metabolic rift of the wooly adelgid-hemlock relationship, we may need to create a metabolic shift within Bacillus subtilis through a genetic rift with Nicotiana tabacum.
Below are the genetic inserts necessary to give Bacillus the requisite tools for nicotine production in a pyridine-pyrrolidine condensation reaction.
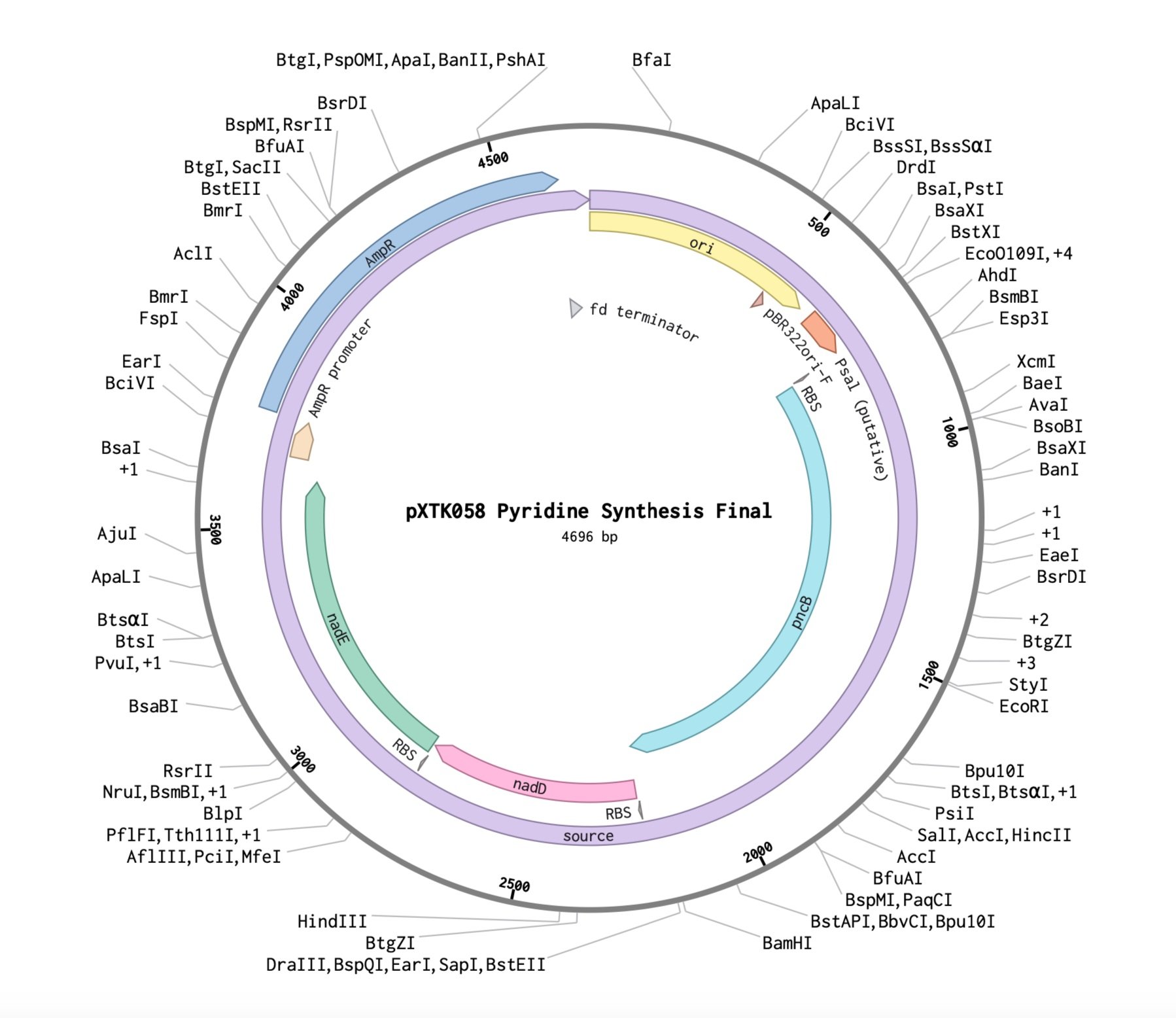

Translating for the Hemlock
I sought to characterize the VOCs emitted by Hemlock during stress, specifically induced during attack by the wooly adelgid. One candidate molecule was methyl salicylate. So, I did research on bacterial protein receptors that might be responsive. I found Pseudomonas putina, a bacterium which has a salicylate-sensitive promoter called Psal. This would be the perfect promoter to have upstream of the nicotine pathway so that nioctine production is controlled and localized to moments of hemlock stress. The image on the right shows a plasmid which contains a salicylate processing enzyme which converts the molecule into an active form that will bind to the Psal promoter and enable transcription. The nicotine plasmids above already have Psal upstream of the enzyme modules.